タコロドプシン光反応過程の研究
<序>
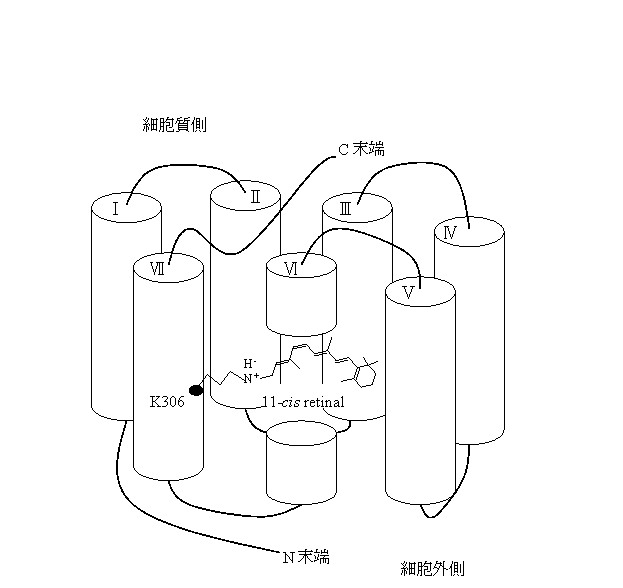
ロドプシン は、動物の視覚システムにおいて最初に光を受ける視覚色素であり、オプシン(タンパク質部分)と11-cisレチナール (発色団)により構成される膜タンパク質である。ロドプシンが光を吸収すると、まずレチナールが11-cisからall-transへ異性化し、その後、熱反応によって吸収極大波長の異なる幾つかの短寿命中間体が生成する。この一つが、Gタンパクと呼ばれるタンパク質と相互作用することにより、視細胞の情報伝達系が駆動し、視神経に視興奮を起こさせる。ロドプシンの光反応過程についての研究は、脊椎動物、特にウシのロドプシンを用いた研究が盛んに行われ、その反応機構はかなり解明されている。一方、無脊椎動物ロドプシンの光反応過程については、比較的まだ不明な点が多い。そこで本研究では、反応によるエネルギー変化、及びタンパク質構造変化をとらえることが可能である、過渡回折格子法(TG法)、過渡レンズ法(TrL法)、光音響信号法(PA法)により、タコロドプシンの光反応過程について、大きく分けて2つの目的を持って研究を行った。
<目的>
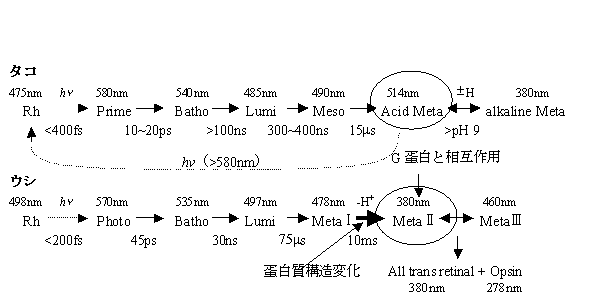
1.脊椎動物の場合、Gタンパクを活性化する中間体が数十ミリ秒のオーダーで、特異的な吸収変化を伴い形成される事が分かっている。一方タコロドプシンの場合、吸収変化は数十マイクロ秒で完了する。しかし、最近、吸収変化完了後も、タンパク質部分だけが構造変化を起こしているのではないかという研究データが得られた。そこで二つ目の目的は、発色団吸収変化をモニターすることでは捉えることができないタンパク質構造変化を、時間分解でとらえることが可能であるTG法により、この構造変化がどの程度の時間オーダーで起き、どの程度の構造変化が起きているか検出することである。
2.エンタルピー及び体積変化は非常に基本的な値であり、ロドプシン光反応過程の理解にとって非常に重要である。しかしこれまでは、反応中間体のエネルギー変化はその中間体が安定に存在できる極低温にトラップする事によって測定されてきた。しかし、極低温と実際の反応の起こる温度では、そのエネルギー変化、構造変化に違いがあるかもしれない。そこで一つ目の目的は、タコロドプシン光反応過程におけるエンタルピー及び、体積変化をTG法、TrL法及びPASにより実際に光反応の起こる条件で求めることである。
<実験>
測定は、色素レーザー(465 nm)を励起光とし、He-Neレーザー(633 nm)及びダイオードレーザー(840 nm)をプローブ光として用いた。タコロドプシンサンプルは、pH7.4 SM1200ミセル溶液に可溶化されたものを用い、濃度は約0.08 mMである。定量測定の際の参照物質としては、励起後直ちに(>1ns)吸収した光エネルギーを全て熱エネルギーとして放出するブロモクレゾールパープル(BCP)を用いた。
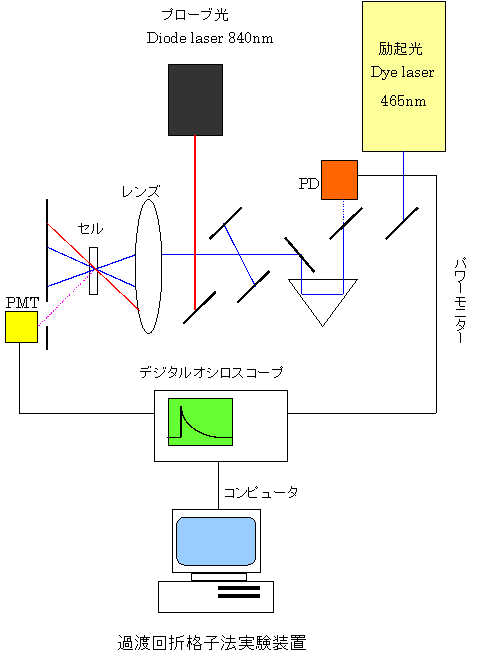
TG、TrL、PASの詳細については他を参照。
<結果・考察>
室温における比較的速い時間スケールでのTG信号
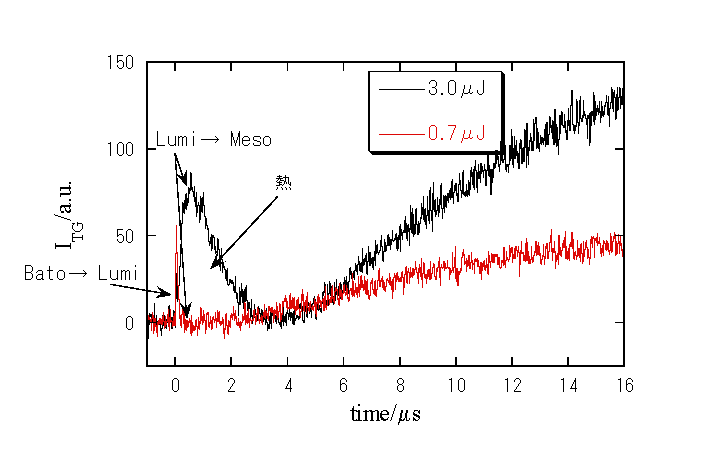
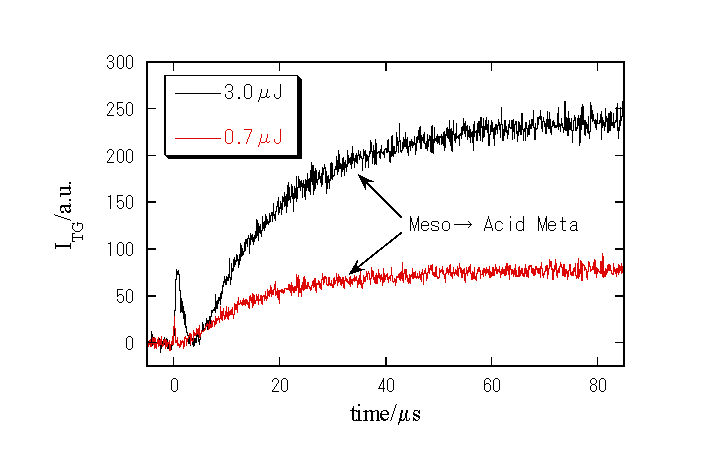
1. 新しい中間体の検出
i) 室温(約23℃)での更に遅い時間領域でのTG信号
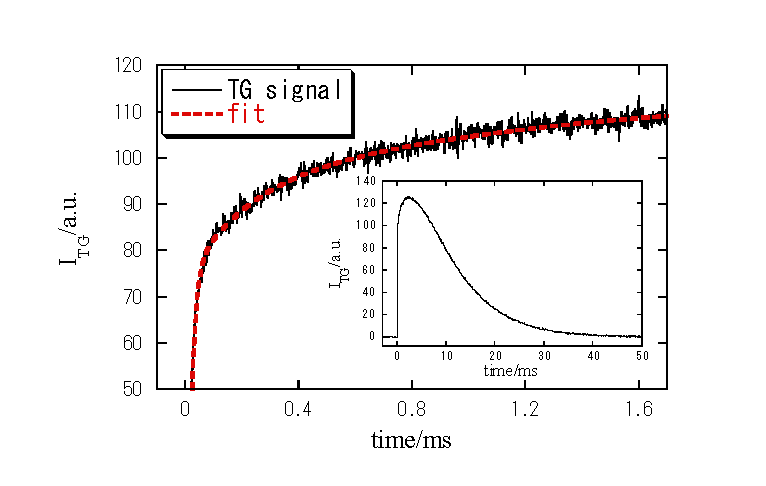
1. 新たな成分の寿命は約150μsであり、Meso→Acid meta及びその後の質量拡散とも異なる。
2.過渡吸収(可視吸収変化)では観測されない。
発色団から離れた部分でのタンパク質構造変化に対応
この構造変化の大きさ13.0 ml/mol
ii) 温度変化測定
新しい成分の温度変化測定を行ったところ、先の吸収変化のダイナミクスとは異なる変化を示した。→発色団から離れた位置での構造変化
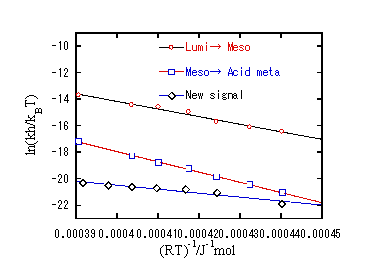
iii) 考察
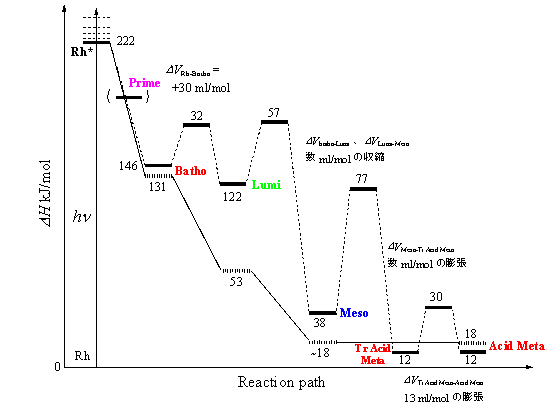
今回観測されたシグナルは、発色団から離れた部分でのタンパク質構造変化、つまりHelixの構造変化に対応していると考えられる。温度変化測定から得られた、活性化エンタルピー、エントロピーの値は、Lumi→Meso→Acid Metaの過程で得られた値と大きく異なる。Lumi→Acid metaの過程は発色団周辺の構造変化を反映しているので、この値と大きく異なることは、観測された構造変化が発色団から離れた位置での構造変化に対応することを支持している。
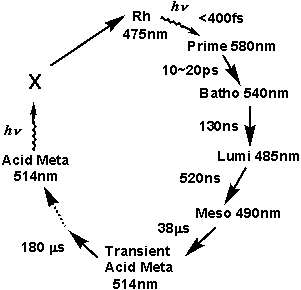
iv)新しいタコロドプシン光反応過程
・発色団の吸収変化の完了に伴い生成する中間体を、Transient Acid Meta。
・今回観測された発色団の吸収変化(発色団周辺の構造変化がない)を伴わず、タンパク質(オプシン部分)のみの構造変化により生成する中間体を一応、Acid Meta。
2.エンタルピー及び体積変化
i) エンタルピー変化
TG信号の熱成分は、Rh*(光励起ロドプシン)からMesoまでの反応で放出された熱による。よって各段階での熱成分の大きさをFittngにより求め、参照物質との比較を取ることにより各中間体のエンタルピー変化を求めることが出来る。
ΔHBatho = 145.77 kJ/mol、ΔHLumi = 121.97kJ/mol、ΔHMeso = 37.6 kJ/molとなる。
MesoからTr Acid Metaの反応での熱はTG信号に観測されないのでTrL法で求めた。TrL法では熱信号がTG法に比べ、比較的長い時間スケールで観測できるためである。
ΔHTr Acid Meta = 12 kJ/molが得られた。
ii)体積変化
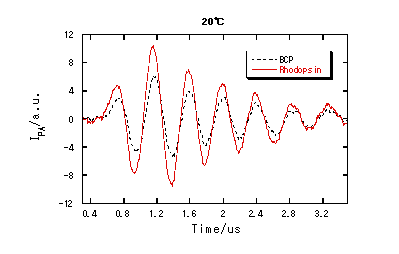
各反応段階での体積変化は、PASによりもとめた。
PASの解析から得られた体積変化は、Rh*からBathoで、比較的大きな約29 ml/molの膨張で、その後の段階では比較的小さな変化しか観測されない。
ii)考察
Bathoは、励起エネルギーの半分以上を蓄えている。このメカニズムは、all-transレチナールの平面からのねじれ、プロトン化シッフ塩基と対イオン間の静電相互作用の変化、及びレチナールの異性化にともなう蛋白部分との立体障害などによる。比較的大きなこの段階での体積変化は、レチナールの異性化に伴う蛋白部分の構造変化によるものと考えられる。
その後の中間体のΔHは徐々に減少するが、これは、先のレチナールのねじれ、立体障害の緩和等によるものと考えられ、この時の変化がΔVとして観測されていると考えられ、この変化は比較的小さいものと考えられる。Lumi中間体で低温とΔHが大きく異なるのは、常温と低温で構造が異なることによる。
Tr Acid Meta生成により吸収変化は完了するので、この時点で発色団周辺の構造変化は完了していると考えられる。この後の吸収変化を伴わない構造変化は比較的大きく13 ml/molであるので、比較的小さな発色団周辺の構造変化の後、比較的大きなHelix部分の構造変化が起きていると考えられる。
以上がタコロドプシンの研究について。
詳しくは、論文を見てください。
A spectrally silent transformation in the photolysis of octopus rhodopsin: a protein conformational change without any accompanying change of the chromophore's absorption.
Yoshinori Nishioku, Masashi Nakagawa, Motoyuki Tsuda, and Masahide Terazima
Biophysical Journal, Vol. 80, pp. 2922-2927 (2001)
その他の研究。。。
修士課程では、脂質の過酸化からの防御反応におけるトンネル効果の研究をという今とは全く違う研究をしてました。
詳しくは、論文を見てください。
Tunneling Effect in Regeneration Reaction of Vitamin E by Ubiquinol.
Shin-ichi Nagaoka, Yoshinori Nishioku, Kazuo Mukai
Chemical Physics Letters, Vol.287, pp. 70-74 (1998)
Tunneling Effect in Antioxidant, Prooxidant, and Regeneration Reactions of Vitamine E.
Shin-ichi Nagaoka, Masayo Inoue, Chiho Nishioka, Yoshinori Nishioku, Sayuri Tsunoda, Chikage Ohguchi, Keishi Ohara, Kazuo Mukai, and Umpei Nagashima
The Journal of Physical Chemistry B, Vol. 104, pp. 856-862 (2000)
Time-Resolved EPR Investigation of the Photo-initiated Intramoleculer Antioxidant Reaction of Vitamin K-Vitamin E Linked Molecule
Yoshinori Nishioku, Keishi Ohara, Kazuo Mukai, and Shin-ichi Nagaoka
The Journal of Physical Chemistry B, Vol. 105, pp. 5032-5038 (2001)