TGチーム研究内容一覧へ
光物理化学研究室へ
☆光回復酵素のDNA光修復反応における蛋白質-DNA間反応ダイナミクス☆
序論
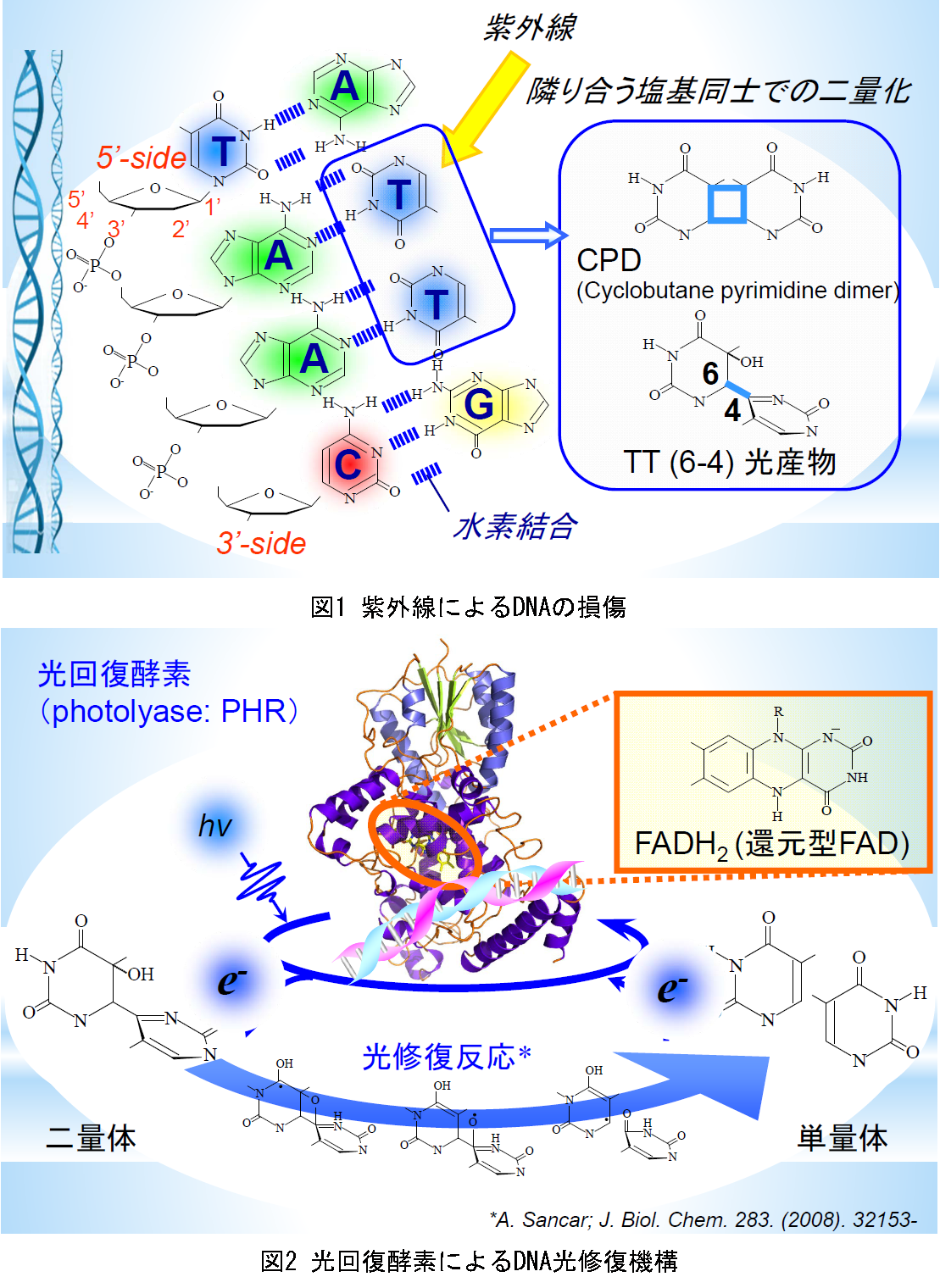 光は生物にとって不可欠である一方で、時に有害な一面も持ちます。その大きな例が紫外線によるDNAの損傷です。DNAは図1に示した様に核酸塩基同士が水素結合により二重鎖を形成し多構造を持ちますが、紫外線の照射によって隣り合う塩基同士の間で二量化反応が起きて、シクロブタンピリミジンダイマー(CPD)や(6-4)光産物((6-4)PP)のような二量体ができます。こうした部分では塩基間の二重鎖ができず、DNAに損傷が起こります。
光は生物にとって不可欠である一方で、時に有害な一面も持ちます。その大きな例が紫外線によるDNAの損傷です。DNAは図1に示した様に核酸塩基同士が水素結合により二重鎖を形成し多構造を持ちますが、紫外線の照射によって隣り合う塩基同士の間で二量化反応が起きて、シクロブタンピリミジンダイマー(CPD)や(6-4)光産物((6-4)PP)のような二量体ができます。こうした部分では塩基間の二重鎖ができず、DNAに損傷が起こります。
こうしたDNAの損傷は放っておくと遺伝的な障害を生み出します。そこで生物はいくつかのDNA修復機構を得てきました。その一つが光回復酵素(PHR)による光修復機構です。PHRは図2に示した様な構造を持ち、発色団として還元型のフラビン(FADH-)を持っています。PHRは暗状態で損傷DNAと結合していて、光誘起により損傷部位に電子を渡した後、二量体を単量体へと戻すことで、DNAを修復することが知られています。しかし、光修復反応過程においてPHRとDNAの間で起こる分子間反応のダイナミクスは全く捉えられてきていません。そこで、本研究ではPHRのDNA修復反応において、PHRとDNA間で起こる分子間反応のダイナミクスを時間分解で捉えることでDNA修復反応の機構を明らかにすることを目指しました。この目的のために、過渡吸収(TA)法と過渡回折格子(TG)法を用い、発色団付近で起きる局所的な電子移動の様な反応から、PHRとDNA全体で起きる反応過程までを時間分解で調べました。(ここではTG測定の結果を紹介します。)
蛋白質とDNAの様な高分子間で起きる反応過程を時間分解で捉えることは従来の手法では困難でした。ここではTG法を用いて分子拡散過程を時間分解で観測するという新しい観点から高分子間反応過程を捉えることを目指しました。酵素には(6-4)PHRという損傷のうち(6-4)PPのみを特異的に修復する酵素を用い、DNAとして配列に一箇所の損傷を含む14塩基対のオリゴヌクレオチドを用いました。負のコントロールとして同配列で損傷を含まないDNAも用いました。PHR、PHR+非損傷DNA、PHR+損傷DNA溶液でTG測定を行い、結果を比較しました。
結果と考察
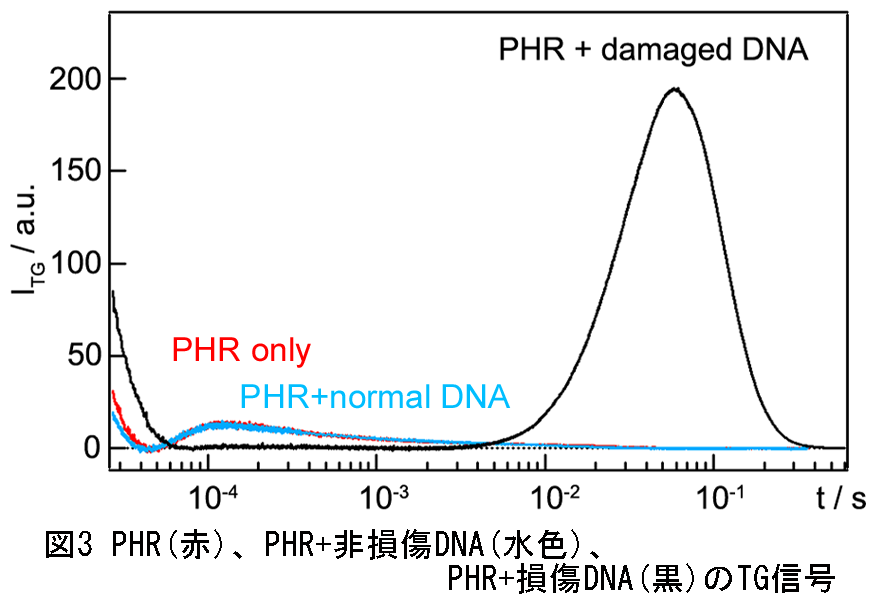 図3にPHR、PHR+非損傷DNA、PHR+損傷DNA溶液の光励起後に観測されたTG信号を示しました。縦軸はTG信号(回折光)の強度、横軸は時間の対数となっています。損傷DNAを加えた時にだけ、(PHRの光励起前と後における)分子拡散過程の変化を反映する特徴的な山型信号が観測されました。このことから、DNA光修復反応過程においては、PHR自身の光反応では見られない様な分子拡散過程の変化が起きることが示されました。
図3にPHR、PHR+非損傷DNA、PHR+損傷DNA溶液の光励起後に観測されたTG信号を示しました。縦軸はTG信号(回折光)の強度、横軸は時間の対数となっています。損傷DNAを加えた時にだけ、(PHRの光励起前と後における)分子拡散過程の変化を反映する特徴的な山型信号が観測されました。このことから、DNA光修復反応過程においては、PHR自身の光反応では見られない様な分子拡散過程の変化が起きることが示されました。
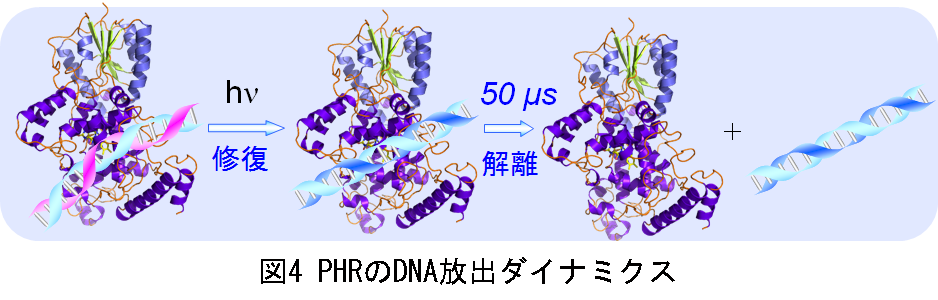
この拡散変化の由来を、分子拡散係数(D)値から評価しました。拡散信号を解析すると、Dの値を得ることができます。この山型の拡散信号を解析すると、この山型信号はPHRに相当するD値を持った化学種(減衰成分)と、これより約2.2倍小さいD値(約10(=2.23)倍小さな分子サイズ)を持った化学種(立ち上がり成分)の拡散過程を反映していることが分かりまし。このことから、観測された分子拡散過程の変化は修復されたDNA (~8 kDa)がPHR-DNA複合体(~ 68 kDa)から溶液中に放出されたという事実、すなわちDNAの解離反応を反映したものと結論しました。格子波数(過渡回折格子の格子間隔)を変えて拡散信号の時間依存性を測定した結果、分子拡散信号を反映する信号強度に時間依存性が観測されました。この時間依存性の解析から、PHRが修復後にDNAを放出する反応の時定数が約50 μs(図4)であると世界で初めて決めることができました。
TGチーム研究内容一覧へ