花の器官形成の遺伝的制御白石英秋(岡崎国立共同研究機構・基礎生物学研究所(執筆当時))
花の器官形成のような高次の生命現象を解析するためには、突然変異体を分離して、正常な花を形づくるためにはどのような遺伝子が必要かを調べることが有効な手段である。花の形態に関する突然変異で特に興味深いのは、花の器官が相互に転換したものが存在することである。生物体を構成するある器官がそれと相同的な別の器官に転換する変異は、ホメオティック変異と呼ばれている(2)。ホメオティック突然変異体は、器官の原基が将来どのような器官になるかを指令する遺伝子が突然変異を起こしているものと考えられ、生物の形態形成のしくみを理解する上できわめて重要な突然変異体である。
【規則2】 突然変異体の器官が変化する方向には、ある種の対称性がある。突然変異体Aは、外側の二つの器官(がく片と花弁)が内側の二つの器官(雄しべと心皮)に鏡像のように対照的に変化している(Fig. 2)。逆に、Cでは、内側の二つの器官(雄しべと心皮)が外側の二つの器官(がく片と花弁)に対照的に変化している。また、B群の突然変異体では、外側から2番目の器官が本来一番外側にあるべき器官に変わり、内側から2番目の器官が本来一番内側にあるべき器官に転換している。この変化のしかたも、ある種の対称性を持っている。
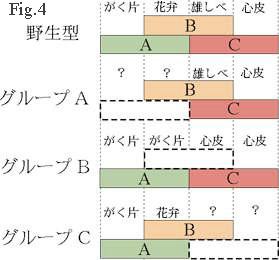
まず、それぞれの遺伝子が、それぞれの突然変異によって異常になる器官で発現しているものと考えると、規則1から、Fig. 4 (左図)の上に示したモデルをつくることができる。このモデルでは、3種類の遺伝子群(A、B、C)がそれぞれ隣り合った2つの領域で発現しており、遺伝子の組み合わせによって、形成される器官が決定されると仮定している。すなわち、遺伝子Aだけが発現している時にがく片ができ、遺伝子AとBが同時に発現すると花弁ができる。また、BとCが同時に発現すると心皮ができると考えている。 このモデルによると、グループBの表現型は、うまく説明することができる。すなわち、この突然変異体では、遺伝子Bの機能が失われたために、外側から2番目の領域で遺伝子Aだけが発現することになってがく片が形成され、同様に、外側から3番目の領域では遺伝子Cだけが発現するために心皮が形成されると説明できる(Fig. 4)。ところが、このモデルでは、AとCの突然変異体の表現型をうまく説明することができない。例えば、グループAの突然変異体の一番外側と外側から2番目の領域では、それぞれ心皮と雄しべが形成されるが(Fig. 2)、モデルでは野生型にみられない遺伝子の組み合わせができてしまうため、何が形成されるか予測不可能である(Fig. 4)。
なお、Fig. 2 に表した表現型は、説明を簡単にするために実際より単純化しており、それぞれの突然変異体の表現型には対立遺伝子ごとに若干の相違がみられる。しかし、それらの相違も、遺伝子AとCの抑制の度合いの違いなどを想定することによってうまく説明することが可能である。 最近、BやCに対応する遺伝子がクローニングされ、それらの遺伝子の構造解析や遺伝子産物の機能の解析から、いずれもDNAの特異的な塩基配列に結合する転写調節因子をコードしていることが示唆されている(4, 5)。この分野の研究は近年著しく進展しており、近い将来には、花の形態形成という植物の一生の中で最も劇的な過程のひとつを、分子レベルの言葉で説明することができるようになるものと期待している。 (1)Coen, E. S. and Meyerowitz,
E. M., Nature, 353, 31-37 (1991). 白石英秋, 花の器官形成の遺伝的制御, Plant Science Tomorrow, NO.6, pp.6-8 (Plant Science Tomorrow 刊行会, 1992)を一部改変。 |
 高等植物の花は一般に高い対称性を持っており、それぞれの植物種に応じて決まった位置に決まった器官が形成される。このことは、花の器官形成に厳密な遺伝的制御機構が存在することを示している。
高等植物の花は一般に高い対称性を持っており、それぞれの植物種に応じて決まった位置に決まった器官が形成される。このことは、花の器官形成に厳密な遺伝的制御機構が存在することを示している。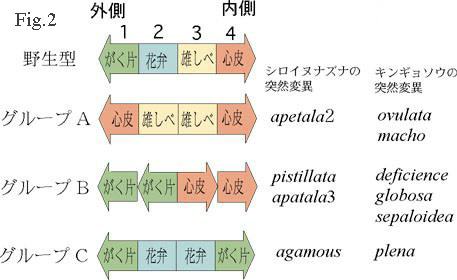 最近、シロイヌナズナ(Fig.
1)とキンギョソウで、花に関する突然変異体の分離と解析がおこなわれ(1,3)、その結果、花のホメオティック突然変異の表現型が、いずれの植物種でも、基本的に3種類に分類できることが明らかになった(Fig.
2)。シロイヌナズナの花は、外側から順番に、4枚のがく片、4枚の花弁、6本の雄しべ、および、2枚の心皮が融合してできた1本の雌しべから成っているが、例えば、突然変異体のひとつ、pistillata
では、花弁ががく片に転換し、雄しべが心皮(雌しべの構成単位)に転換しているため、外側から順番にがく片、がく片、心皮、心皮というように器官が形成される(Fig.
2 、グループB)。Fig. 3は、この pistillata 突然変異体の花で、6本の雄しべが、すべて心皮(雌しべの構成単位)に変わっている(この写真では、一番外側のがく片と、その内側の花弁が変化してできたがく片は取り除いてある)。さて、Fig.
2の突然変異体の表現型をよく見ると、次のような規則性があることがわかる。
最近、シロイヌナズナ(Fig.
1)とキンギョソウで、花に関する突然変異体の分離と解析がおこなわれ(1,3)、その結果、花のホメオティック突然変異の表現型が、いずれの植物種でも、基本的に3種類に分類できることが明らかになった(Fig.
2)。シロイヌナズナの花は、外側から順番に、4枚のがく片、4枚の花弁、6本の雄しべ、および、2枚の心皮が融合してできた1本の雌しべから成っているが、例えば、突然変異体のひとつ、pistillata
では、花弁ががく片に転換し、雄しべが心皮(雌しべの構成単位)に転換しているため、外側から順番にがく片、がく片、心皮、心皮というように器官が形成される(Fig.
2 、グループB)。Fig. 3は、この pistillata 突然変異体の花で、6本の雄しべが、すべて心皮(雌しべの構成単位)に変わっている(この写真では、一番外側のがく片と、その内側の花弁が変化してできたがく片は取り除いてある)。さて、Fig.
2の突然変異体の表現型をよく見ると、次のような規則性があることがわかる。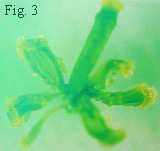 【規則1】 それぞれの突然変異体は、単一突然変異によって、となりあった二つの器官が同時に別の器官に転換している。すなわち、突然変異体Aではがく片と花弁、突然変異体Bでは花弁と雄しべ、突然変異体Cでは雌しべと心皮がそれぞれ別の器官に変化している(Fig.
2)。
【規則1】 それぞれの突然変異体は、単一突然変異によって、となりあった二つの器官が同時に別の器官に転換している。すなわち、突然変異体Aではがく片と花弁、突然変異体Bでは花弁と雄しべ、突然変異体Cでは雌しべと心皮がそれぞれ別の器官に変化している(Fig.
2)。 表現型にこのような規則性があるという事実は、遺伝子の作用のしかたを反映しているものと考えられよう。表現型の規則性から、どのような遺伝子作用のモデルが考えられるだろうか。
表現型にこのような規則性があるという事実は、遺伝子の作用のしかたを反映しているものと考えられよう。表現型の規則性から、どのような遺伝子作用のモデルが考えられるだろうか。 ここで、規則2を考慮に入れて、突然変異体(特にグループAとCに属するもの)での遺伝子発現が対称性を持つようにモデルを改変するとFig.
5(左図)のようになる。このモデルでは遺伝子Aと遺伝子Cが中央部で互いの発現を抑制しあっており、遺伝子Aまたは遺伝子Cの機能が失われると、それぞれ遺伝子C、Aが花芽全体で発現すると仮定している。このモデルでは、遺伝子の組み合わせからそれぞれに部位に形成されると予測される器官が、実際の表現型と完全に一致している。また、この改良されたモデルでは、二重突然変異体の表現型もうまく予測することができ、それは実際の二重突然変異体の表現型と一致する。
ここで、規則2を考慮に入れて、突然変異体(特にグループAとCに属するもの)での遺伝子発現が対称性を持つようにモデルを改変するとFig.
5(左図)のようになる。このモデルでは遺伝子Aと遺伝子Cが中央部で互いの発現を抑制しあっており、遺伝子Aまたは遺伝子Cの機能が失われると、それぞれ遺伝子C、Aが花芽全体で発現すると仮定している。このモデルでは、遺伝子の組み合わせからそれぞれに部位に形成されると予測される器官が、実際の表現型と完全に一致している。また、この改良されたモデルでは、二重突然変異体の表現型もうまく予測することができ、それは実際の二重突然変異体の表現型と一致する。